В отличие от соматических клеток многоклеточных протисты (клетка-организм) характеризуются наличием жизненного цикла, который слагается из ряда следующих друг за другом стадий, повторяющихся у каждого вида с определенной закономерностью (Догель, 1981).
Цикл развития, или жизненный цикл вида лежит в основе его полного морфологического описания, ибо под формой организма понимается весь цикл его изменений (морфопроцессов) в онтогенезе (Беклемишев, 1994). Существует несколько определений этого процесса, суть которых сводится к следующему. Жизненным циклом называется совокупность всех стадий или фаз развития, которые последовательно проходит организм любого вида, начиная и заканчивая одними и теми же моментами, например, от зиготы до зиготы. Этим же термином называют цитологическую и морфологическую последовательность событий, происходящих при половом воспроизведении, а также более широкое понятие, которое может быть определено как сумма адаптаций организма к выживанию и размножению (Саут, Уиттик, 1990). Протяженность жизненного цикла определяется числом поколений (генераций), развивающихся в течение года или времени, в течение которого осуществляется один жизненный цикл. Длительность его зависит от продолжительности обязательного для организма периода покоя или диапаузы (Биологический энциклопедический словарь, 1989). В цикле развития любого вида, в том числе динофитовых водорослей, период покоя есть, вероятно, обязательный этап.
Жизненный цикл динофлагеллят характеризуется в целом зиготическим типом мейоза, который считается наиболее примитивным. Зиготический мейоз происходит в зиготе перед ее прорастанием, поэтому она является единственной диплоидной фазой у подавляющего большинства изученных динофитовых, за исключением ночесветки (Noctiluca), у которой наблюдается гаметический мейоз. Зиготические жизненные циклы чаще всего встречаются у одноклеточных динофлагеллят, а также среди их колониальных и пальмеллоидных представителей.
Жизненные циклы разной степени сложности пока известны у 10% современных видов динофлагеллят (Dale, 1982, цит. по Anderson et al., 1985), что может составлять в настоящее время примерно 200-250 видов. Главным образом они изучены у неритических перидиней. Известны простые и сложные (большие) циклы развития. Последние включают как половой, так и различные типы бесполого размножения. Простые циклы встречаются наиболее часто у агамных свободноживущих планктонных и бенто-планктонных видов, претерпевающих в цикле развития смену морфологически различающихся индивидов, которая носит название цикломорфоза. Последний не нужно путать со сменой форм существования, которая определяется только внешними условиями, т.е. экологией, и не носит циклический характер. Явление адаптивного полиморфизма существует независимо от наличия или отсутствия полового процесса (Петров, 1986). Простые жизненные циклы включают обычно фазы развития только одной особи и имеют, как правило, не более 5-10 стадий (рис. 5). Сложные циклы развития, наиболее часто идущие с метаморфозом, характеризуются чередованием различных типов полового и бесполого размножения и могут иметь до 25-30 стадий. Примером последних могут служить жизненные циклы токсичных представителей Pfiesteria. Виды этого рода имеют большой диапазон размеров (от 5 до 750 мкм) и форм (в 22-24 стадиях) – от мелких бесцветных округлых клеток до крупных многоформных амебоидов в зависимости от типа жертвы и условий окружающей среды (Burkholder, 1999). Относительно сложные циклы развития с метаморфозом присущи и многим паразитическим динофлагеллятам (табл. XXV, 3а-3и). Неритические виды динофитовых имеют, как правило, годовой (зимне-летний) бенто-планктонный цикл с чередованием неподвижной бентической покоящейся споры (цисты) и планктонной двужгутиковой клетки (рис. 6). Жизненный цикл настоящих океанических видов, не имеющих бентосных покоящихся стадий в виде спор, вероятно полностью проходит в пелагиали, где в роли покоящейся диплоидной стадии выступает планозигота.
Впервые исчерпывающая информация относительно развития стадий тека – спора примерно у 30 видов морских планктонных динофлагеллят была получена в конце 60-х годов Воллом и Дали (Wall, Dale, 1968). Свои наблюдения в культурах относительно развития текальных неритических перидиней они обобщили и дали в виде рисунка (Wall, Dale, 1968:text-figure 2), используя в качестве примера Gonyaulax digitalis. Этот цикл, как писали авторы, по-видимому, включает только относительно простые годовые чередования подвижной двужгутиковой клетки, обычно защищенной целлюлозной оболочкой (стенкой), с неподвижной бентосной клеткой – спорой. Существенно то, что авторы показали присутствие еще одной стадии – промежуточной гимнодиниеподобной фазы, образующейся немедленно после эксцистирования, и часто не заметной для исследователей в связи с кратковременностью ее существования. Это тонкостенная, с одним продольным жгутом клетка, которая имеет мягкий бесцветный желатинированный футляр и выходит из споры через археопиль амебоидным движением. В течение нескольких минут становится заметным второй (поперечный) жгутик. Эта фаза, перед образованием панциря, короткая, так как тека развивается в пределах одного дня. Окончательное формирование прочного панциря завершается в течение нескольких дней. Сразу после восстановления подвижной стадии в отдельных случаях наблюдалось быстрое деление клетки.
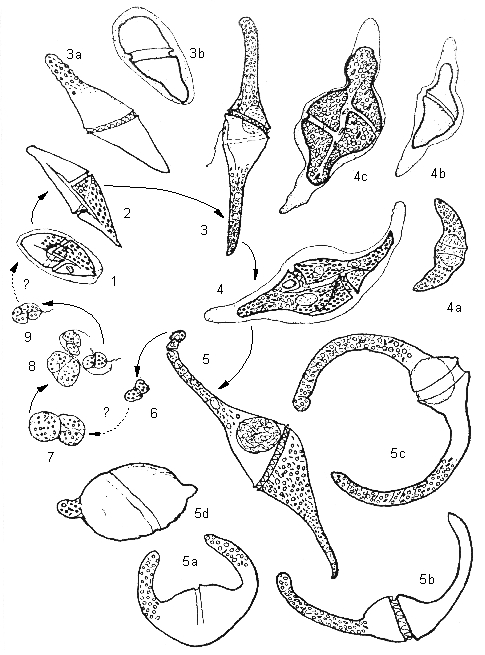
Рисунок 5. Пример относительно простого цикла развития динофлагеллят (жизненный циклGyrodinium falcatum по: Коновалова, 2003).
1, 2 – молодые клетки спереди (вентрально); 3, 3а, 3б – растущие вегетативные клетки: 3 – латерально-вентральный вид клетки в нормальных условиях обитания, 3а – клетка, подверженная тепловому шоку, 3б – клетка под воздействием резкого опреснения воды; 4, 4а–4в – взрослые вегетативные клетки в следующей фазе роста: 4, 4в – вентральный и латеральный (4а, 4б) вид живых (4, 4а) и мертвых клеток (4б, 4в – фиксированные клетки, по: Schütt, 1895); 5, 5а-5г – зрелые организмы (споронты?) в разном положении: 5, 5б – живые клетки в дорсальном и латеральном положении, 5а, 5в – фиксированные клетки, 5г – умирающая клетка (5) после «сбрасывания» апикальных члеников и подверженная сильному опреснению; 6 – «клетки», отчленившиеся от апикальной части зрелого организма; 7 – две подобные, но большего размера клетки с контурами бинарного деления внутри; 8 – разрыв оболочки одной из них и выход дочерних клеток; 9 – жгутиковая клетка, подобная ювенильным стадиям G. falcatum.
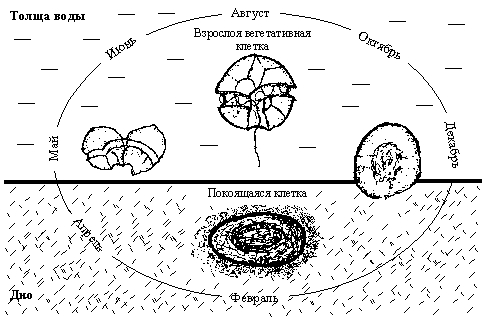
Рисунок 6. Схема упрощенного сезонного цикла развития видов рода Alexandrium на примереA. tamarense (по: Коновалова, 1999).
Споры. Спорами (гр. spora – семя) жгутиковых водорослей называют одноклеточные неподвижные образования (организмы), покрытые в большинстве случаев плотными оболочками и служащие для сохранения размножения и распространения вида. Динофитовые образуют два основных типа спор: временные и покоящиеся споры (цисты). Необходимо заметить, что в иностранной, главным образом англоязычной литературе, все споры на сегодняшний день именуются цистами (гр. kystis – пузырь), термином более уместным для палинологии, откуда он и пришел в необотанику, и применяются в качестве синонима спор, что вряд ли можно считать правильным во всех случаях, так как цистами у динофлагеллят могут быть названы клетки нескольких стадий, несущих разные функции (Fensome et al., 1993), в том числе не относящихся к размножению, например пищеварительные (digestion cyst), т. е. любые пузыревидные или мешковидные образования. В русскоязычной ботанической литературе цистой называется покоящаяся спора низших растений (жгутиковых водорослей и низших грибов), характеризующаяся толстыми прочными оболочками и служащая для перенесения неблагоприятных условий (Словарь бот. терм., 1984). Поэтому термин «циста» в русскоязычной литературе следует применять с оговоркой.
Временные споры образуются, как правило, в результате отчетливо меняющихся условий среды обитания. Это могут быть резкие изменения температуры и солености, пищевое голодание, изменение химизма среды в результате накопления метаболитов (например, в культурах), а также механические стрессы при сильном движении воды или чрезмерном скоплении организмов. При этом подвижные клетки часто сбрасывают жгуты и покрываются тонкой и мягкой, но достаточно прочной прозрачной оболочкой, внутри которой тело нередко округляется, хотя может оставаться и без изменений (рис. 7). У текальных видов при стрессах иногда наблюдается сбрасывание панциря при сохранении жгутов, при этом голая гимнодиниеподобная клетка сохраняет способность к движению (см. Коновалова, 2006: рис. 7). Типизация, т. е. приобретение характерного вида временных спор, а также существование и восстановление прежнего облика подвижной клетки, происходит обычно довольно быстро при реконструкции среды – от нескольких минут или часов до нескольких дней, реже недель, что является отличительной способностью этого типа спор.
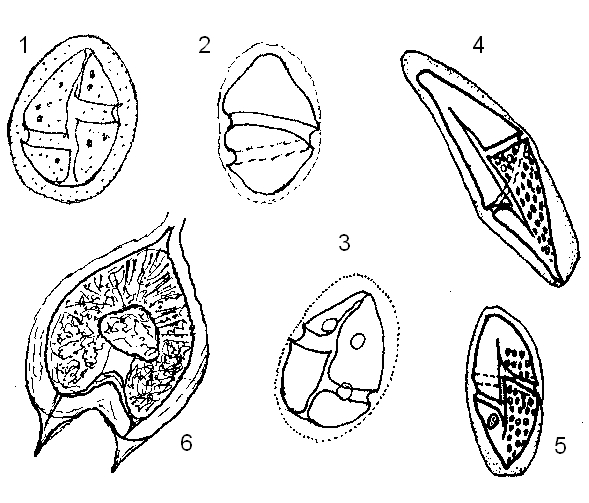
Рисунок 7. Временные споры в пузыревидных цистах с прозрачной оболочкой: 1-5 –Gyrodinium; 6 – Protoperidinium.
Покоящиеся споры, или цисты бывают у динофлагеллят двух типов: бесполые или вегетативные автоспоры (гипноспоры) и половые зигоспоры (гипнозиготы). Циста может быть как внутриклеточной, так и внеклеточной. В первом случае стенка цисты может быть образована из оболочки (панциря, амфиесмы) самой клетки, что сближает ее с акинетой, которая представляет собой модифицированную клетку с утолщенной стенкой. В другом случае оболочка покоящейся споры образуется заново цитопластом, который предварительно отделяется от стенок родительской клетки. Она может быть укреплена устойчивым материалом типа спорополленина (Dodge, Grevet, 1987, цит. по: Taylor, 1987a). Такие споры могут сохранять жизнеспособность в природе в течение многих лет, а в лабораторных условиях даже до 20 лет (Coleman, 1983, цит. по: Саут, Уиттик, 1990). Чаще всего покоящиеся споры динофлагеллят представляют собою зиготы, т. е. клетки, которые являются продуктами полового размножения. Циста-зигота имеет несколько преимуществ перед обычной гаплоидной спорой. Она обладает большой генетической изменчивостью, устойчивостью к неблагоприятным условиям среды обитания, но главным образом служит средством размножения (Dale, 1983). Покоящиеся споры образуются в ответ на резко меняющиеся условия среды (понижение температуры, пищевое голодание и др.), но в основном при завершении естественной стадии жизненного цикла или в конце вегетационного периода. У текальных динофлагеллят они формируются, как правило, эндогенно в любое время года, но обычно ассоциируются с «цветением», концом вегетации или после периода экспоненциального роста. Наибольшая численность цист в морях приурочена к осени. При этом клетки теряют подвижность и, оседая, уходят из планктона. Дезинтеграция материнской теки происходит по-разному. Так, у видов рода Gonyaulaxпанцирь разрушается быстро, а у Protoperidinium этот процесс может длиться много дней (Wall, Dale, 1968). Замечено, что у некоторых видов последнего рода материнская тека в процессе спорообразования может изменяться, приобретая некоторые черты оболочки споры (рис. 8, 13). Часть цист динофитовых заканчивает спецификацию (становление зрелого морфотипа цисты) уже в толще воды, оседая и избавляясь от материнской теки. Очевидно это происходит главным образом у океанических видов, обитающих над большими глубинами (Akselman, 1987). Отмечено, что цисты перидиней очень редко встречаются в современных донных океанических осадках (Жузе, 1962; Коренева, 1964, цит. по: Возженникова, 1967). Неритические виды, как правило, завершают спецификацию на дне. В любом случае цисты, достигшие дна, уже, как правило, не имеют остатков материнского панциря, о чем свидетельствуют исследования Матсуоки (Matsuoka, 1985а, 1987). Зрелые цисты могут напоминать вегетативные клетки, например у Protoperidinium, или сильно отличаться от них (Gonyaulax). Их оболочка (стенка или фрагма) в большинстве случаев укреплена спорополленином, надежно защищающим организм от агрессии окружающей среды, что является характерной особенностью покоящихся спор, в особенности зигоспор. Содержимое этих цист включает пищевые резервы, такие как крахмал и масло, часто пигментированный «глазок» (красное пятно?), ядро и микрогранулы цитоплазмы. Споры, достигшие дна (а над большими глубинами, вероятно, и в толще воды), должны пройти обязательную стадию внутреннего созревания прежде, чем они смогут прорасти при наличии благоприятных условий. У разных видов она длится приблизительно от 6 недель до 9 месяцев. Матсуока и Фукуё (Matsuoka, Fukuyo, 2000) выделяют две формы покоящихся спор, существующих в верхних слоях современных осадков: живые, т.е. споры, содержащие зародыши, способные прорасти при благоприятных условиях, и мертвые споры, т.е. пустые оболочки, оставленные после выхода протопласта (эксцистирования) через специальное отверстие – археопиль. Внешний вид цисты одного и того же вида в процессе созревания и деградации ее оболочки после эксцистирования может меняться, и живые споры, найденные в планктоне зачастую имеют несколько другие морфологические характеристики, чем мертвые цисты, лежащие на дне. Однако обнаружить зрелые цисты в пелагиали гораздо труднее из-за малого их количества, т.к. только одна из нескольких сотен подвижных клеток динофлагеллят способна их продуцировать (Matsuoka, Fukuyo, 1987). В морях и океанах споры динофитовых встречаются до глубины 500 м, но наиболее часто – в верхнем 30-метровом слое воды. Вероятно, большое количество покоящихся спор в донных осадках накапливается в течение многих лет. Наибольшая концентрация их в планктоне, как правило, отражается на высокой численности – в осадках, но это бывает не всегда, вследствие перемещения цист течениями от берегов в более мористую зону (Reid, 1978). Концентрация покоящихся спор в верхних слоях донных отложений в морях может достигать 200-300 и даже более 1000 экз./см3(Nehring, 1995), однако, как показали исследования Кобаяси и Юки (Kobayashi, Yuki, 1991), около 60-88% их представлены пустыми цистами, т.е. одними оболочками. Покоящиеся споры в жизненном цикле пока известны более чем у 10% современных видов динофитовых (Head, 1996), что на сегодняшний день может составлять около 200-250 панцирных и беспанцирных видов. Причем споры последних могут быть более многочисленными в морских поверхностных отложениях. Однако известно, что они часто имеют слабые, не устойчивые к агрессии морской воды оболочки, быстро разрушаются и не выносят фоссилизации. Это свидетельствует о том, что химически они менее устойчивы, чем споры текальных видов, образующих типичные гистрихосферы – ископаемые цисты панцирных динофлагеллят (Wall, Dale, 1968; Blanco, 1987).
Значительное число современных панцирных динофитовых образуют покоящиеся споры, которые имеют непосредственное палеонтологическое значение. Сюда относятся главным образом роды: Gonyaulax, Protoperidinium, Protoceratium. В современных осадках неритической зоны морей, помимо перечисленных, часто встречаются цисты Alexandrium,Scrippsiella, Diplopsalis, Pyrodinium, Lingulodinium, Gymnodinium, Amphidinium и др. (рис. 8) Сравнительно недавно обнаружены споры пророцентроидов и динофизоидов.
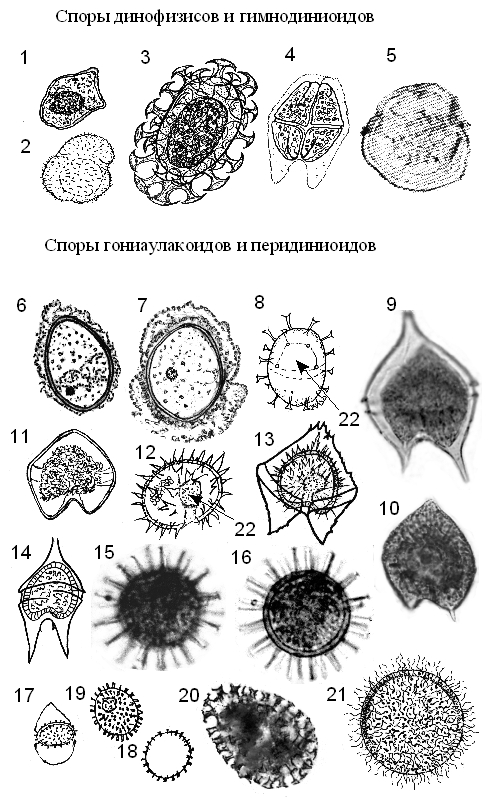
Рисунок 8. Наиболее распространенные типы покоящихся спор (цист): 1 –Dinophysis sp.; 2 –Amphidinium larvale; 3 – Polykrikos schwartzii; 4 – Akashiwo sanguinea; 5 – Gyrodiniuminstriatum; 6 – Gonyaulax verior; 7 – Alexandrium sp.; 8 – Gonyaulax spinifera (из Коновалова, 1998 по: Dodge, 1985a); 9, 10 – Protopoperidinium claudicans; 11 – P. oblongum; 12, 13 – P.conicum; 14 – P. oceanicum; 15, 16 – Protoceratium reticulatum; 17-20 – Scripppsiella trochidea(20 – S. cristalina?); 21 – Protoperidinium minutum; арх. – археопиль.
Причем, единичные споры (цисты) Prorocentrum marinum и P. lima (Faust, 1990a, 1993b), а также стадии планозиготы и цисты Dinophysis accuminata, D. acuta и D. tripos (Mc Laehlan, 1993; Moita, Sampayo, 1993) найдены только в толще воды и никогда не были обнаружены в осадках. Некоторые исследователи (Matsuoka, Fukuyo, 2000) полагают, что споры эти вероятно не сохраняемые. Однако можно предположить, что для большинства пророцентроидно-динофизоидной группы видов, как и для видов таких типично океанических родов (существующих в относительно более стабильных условиях) как Ceratium, Ceratocorys,Heterodinium, Histioneis, Ornithocercus и Oxytoxum, у которых не найдены цисты (или очень редко), не характерны покоящиеся бентические стадии в виде спор, и весь цикл развития происходит в толще воды, где роль зиготы выполняет планозигота. Последняя, как известно, ничем, кроме размеров, существенно не отличается от вегетативных клеток, причисляется обычно к их числу при рутинных исследованиях фитопланктона, и может быть выявлена только при изучении цикла развития вида (Stosch, 1972, цит. по: Петров, 1986). Большинство видов, образующих бентические споры, являются умеренными или эвритермными неритическими, существующими в нестабильной среде, с особенно резко меняющимися температурной и соленостью воды. Механизм образования покоящихся спор, как пишут Волл и Дэйл (Wall, Dale, 1968), по-видимому, является высоко адаптивным методом распространения вида. Он может способствовать временной (сезонной) и географической (локальной) изоляции популяций динофлагеллят, поскольку как инцистирование, так и эксцистирование наблюдается в крайне жестко определенных условиях окружающей среды.
Морфологические признаки покоящейся споры, или цисты как заключительной стадии жизненного цикла вида могут быть использованы в систематике для идентификации голоморфа, в особенности у текальных видов. Выделяют стабильные и ограниченные признаки (Wall, Dale, 1968). К стабильным признакам относятся положение и форма археопиля и отраженная табуляция. Второй признак уступает по значимости, так как, если археопиль есть характерная особенность всех цист динофитовых, то отраженная табуляция касается только спор панцирных перидиней и может быть неразвитой у многих видов, например у Protoperidinium. Положение археопиля, как отверстия для эксцистирования в стенке цисты, оказалось абсолютно неизменным, в особенности для современных текальных видов. По характеру совпадения археопиля с местонахождением одной или нескольких пластинок исходной теки выделяют следующие типы археопиля: апикальный, пресингулярный, гипоцистальный и комбинированный, например, апикально-интеркалярно-пресингулярный (см. Matsuoka et. al., 1989; Matsuoka, Fukuyo, 2000: fig. 15). Для беспанцирных и некоторых панцирных видов Матсуока (Matsuoka, 1985b) выделил еще два типа археопиля: чесмик («chasmic», англ. chasm – расселина, трещина), обладающий линейной или слегка изогнутой трещиной, без крышечки (оперкулюма) и расположенный в средней части цисты, и тремик («tremic», гр. trema – отверстие), имеющий округлое отверстие, часто с неровными краями, расположенное на апикальном участке покоящейся споры.
Все вышеизложенные типы археопилей выделяются в три крупные группы, каждая из которых имеет соответственно сафопильный (saphopylic), теропильный (theropylic) или криптопильный (cryptopylic) археопиль (см. Matsuoka, Fukuyo, 2000: fig. 15). Сафопильный археопиль характерен для цист текальных динофлагеллят порядков Peridiniales иGonyaulacales. Его положение обычно точно совпадает с местоположением одной или нескольких пластинок исходной теки. Теропильный археопиль типичен для цист диплопсалид (Diplopsalis, Diplopelta и др.) и некоторых близких родов. В этом случае швы, ограничивающие археопиль, могут точно не совпадать с положением соответствующих швов панциря. Криптопильный археопиль свойственен покоящимся спорам как текальных, так и, главным образом, атекальных видов динофитовых. Сюда относятся такие типы археопилей, как чесмик и тремик, которые не имеют никаких границ с пластинками у панцирных видов, а у беспанцирных располагаются до некоторой степени произвольно. Археопиль этой группы имеют в основном споры видов Alexandrium, Ensiculifera, Scrippsiella, некоторых Peridinium и др. Из гимнодиниоидов – Gymnodinium, Polykrikos, Cochlodinium, Gyrodinium, Pheopolykrikosи, возможно, Amphidinium.
Отраженная табуляция как термин применяется в тех случаях, когда табуляция паратеки цисты повторяет или напоминает образец табуляции теки родительской клетки. Современные виды с хорошо развитой отраженной табуляцией, т.е. с четким согласием между рисунком стенок цисты и теки, принадлежат к родам Gonyaulax, Peridinium,Diplopsalopsis и Diplopsalis. Однако некоторые цисты современных видов часто не имеют отраженной табуляции, либо имеют лишь неполное повторение табуляции теки вегетативной клетки. К ним относятся цисты видов Protoperidinium, которые редко обнаруживают какие-либо другие признаки строения родительской теки, кроме складок вдоль экваториальной полосы, указывающих на поясок и вентральное углубление борозды, но главное – пятиугольной формы тела (Wall, Dale, 1968).
К другим признакам, которые могут быть использованы как достаточно стабильные, последние авторы относят форму тела и размер спор. Они представляют собой параметры, которые сильно изменяются от вида к виду, но остаются неизменными внутри вида. Так, цисты многих видов Protoperidinium значительно различаются по форме, но споры одного вида жестко сохраняют свою форму. Напротив, цисты Gonyaulax обнаруживают весьма незначительные внутривидовые различия. К нестабильным признакам, варьирующим внутри вида, относятся длина апикального и антапикальных рогов у спор некоторых Protoperidiniumи высота элементов орнаментации у отдельных Gonyaulax и Protoceratium.
Изучение покоящихся спор (цист) динофитовых в дальневосточных морях, в том числе Японском, ранее проводилось лишь эпизодически вместе с исследованием фитопланктона. Первые споры, отнесенные к определенным видам, появились в наших работах (Коновалова и др., 1989; Коновалова, 1998), хотя несколько спор, не идентифицированных ближе рода материнской клетки, были изображены в работах Г. И. Гайла (1950) и И. А. Киселева (1950). Некоторые результаты изучения цист видов рода Alexandrium были коротко представлены в работах конца прошлого века (Коновалова, 1999). Отдельные цисты стали известны при изучении морфологии и жизненных циклов нескольких видов динофлагеллят Японского моря (Коновалова, 2003; Орлова и др., 2003; Орлова, Морозова, 2003). Однако наиболее полная информация относительно их количества получена недавно Т. Ю. Орловой с соавторами (Orlova et al., 2004) при изучении покоящихся спор динофитовых водорослей верхней части донных осадков неритической зоны дальневосточных морей России.
Подробности морфологического строения спор, методы исследования, а также таблицы для их определения и словарь терминов можно найти в работах Матсуоки с соавторами (Matsuoka et al., 1989), Лентина и Возженниковой (Lentin, Vozzhennikova, 1990) и Матсуоки и Фукуи (Matsuoka, Fukuyo, 2000).
Основные работы общей части:
Коновалова, 1998; Lentin, Vozzhennikova, 1990; Matsuoka, Fukuyo, 2000; Taylor, 2004; Wall,Dale, 1968.