Морфология динофитовых достаточно детально описана ранее (Коновалова, 1998), поэтому здесь мы остановимся только на тех основных элементах внешнего строения жгутиковой клетки, которые необходимы для пользования определительными таблицами, поскольку диагностика пелагических и бентических перидиней ведется в основном по строению свободноживущей вегетативной жгутиковой клетки (стадии). Здесь же, насколько возможно полно, постараемся дать информацию о циклах развития динофлагеллят, включая спорообразование и описание спор, так как знание жизненного цикла имеет, на наш взгляд, решающее значение при идентификации вида у перидиней, в особенности при описании беспанцирных и новых для науки видов (Коновалова, 2004).
Динофитовые водоросли (динофлагелляты, или перидинеи) – микроскопические (от 4 до 2000 мкм) жгутиковые, главным образом пелагические одноклеточные протисты округлой, дисковидной или ромбовидной формы, многие из которых несут в разной степени отчетливые выросты на поверхности тела. В онтогенезе или цикле развития флагеллят нередко присутствуют неподвижные, лишенные жгутов клетки, поэтому под жгутиковыми водорослями принято понимать монады, способные размножаться в жгутиковой стадии, независимо от того, имеется или нет безжгутиковая стадия в цикле их развития. Подавляющее большинство динофлагеллят имеет две хорошо выраженные борозды, один из главных фенотипических признаков: продольную (собственно борозду, или сулькус) и поперечную (поясок, или сингулюм) и два гетерогенных жгутика, по типу прикрепления которых они делятся на две основные группы (рис. 2, 3, 4). У наиболее многочисленных – диноконтов – жгуты выходят на брюшной стороне, примерно в середине клетки, жгуты десмоконтов располагаются вблизи верхнего конца клетки.
По строению клеточного покрова (амфиесмы) динофитовые условно делятся на виды, содержащие панцирь (теку), то есть панцирные, или текальные и не содержащие его, то есть беспанцирные, или атекальные, так называемые «голые». Панцирные составляют большинство морских динофитовых водорослей, хотя некоторые из них могут не содержать теки на отдельных стадиях цикла развития. Пластинки панциря, составляющие формулу теки, лежат в основе таксономии и используются как надежные признаки при идентификации динофлагеллят на уровне родов и видов, т.к. положение, количество и взаимная сваязь пластинок в панцире зрелой клетки-организма строго закономерны, имея, как установлено для современных перидиней, систематическое значение (рис. 2). В простейших случаях диагностики сингулярный и особенно сулькальный, как наиболее трудный, анализы могут быть опущены, однако в сложных положениях, а также в ряде случаев при определении видов, например рода Alexandrium, последний необходим. При идентификации видов родаProrocentrum стал широко использоваться порядок расположения и число пор на поверхности теки (Faust, 1990b).
Современные панцирные перидинеи подразделяются по строению теки типового или наиболее многочисленного рода (дан в скобках) на пять основных типов табуляции: пророцентроиды (Prorocentrum), динофизоиды (Dinophysis), гониаулакоиды (Gonyaulax), перидиниоиды (Peridinium, Protoperidinium) и суиссиоиды. Последние включают в себя главным образом ископаемые формы, но имеют и живых представителей родовSymbiodinium и Polarella (см. Taylor, 2004). Пророцентроиды очень похожи на динофизоидов билатеральным строением тела и латеральным делением сагитальным (мегацитарным) швом, но у них, в отличие от динофизоидов, отсутствует борозда и явно выраженный поясок (см. Коновалова, 1998). Кроме того, динофизоиды имеют эпитекальные, поясковые и бороздковые текальные пластинки вокруг жгутов, правда, последние довольно мелкие и трудно обнаруживаемые (рис. 2). Перидиниоиды и гониолакоиды имеют по шесть серий текальных пластинок, но различных по симметрии (рис. 3).
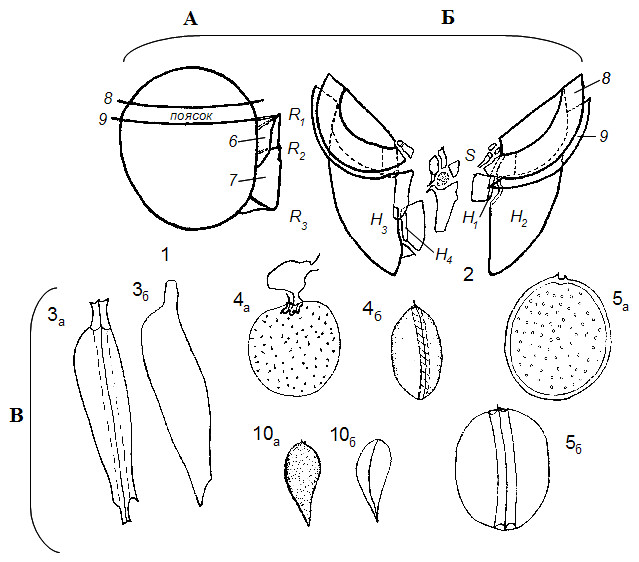
Рисунок 2. Общий вид и схема строения теки основных типов панцирных перидиней: пророцентроиды (В) и динофизоиды (А, Б из: Balech, 1980): 1 – вид клетки Dinophysis cправого бока; 2 – строение теки с брюшной стороны; 3а, 4б, 5б, 10б – вид клетки Prorocentrumcпереди (со шва) и сбоку (со створки) – 3б, 4а, 5а, 10а; 6, 7 – правое и левое крылья борозды; R1 – R3 – ребра большого левого крыла борозды; 8, 9 – верхнее и нижнее крылья (мембраны) пояска; S – пластинки борозды; H1 – H4 – пластинки гипотеки.
Беспанцирные динофитовые называются еще гимнодиниоидами по наиболее распространенному в морях типовому роду Gymnodinium (рис. 4). Их таксономия представляет особую сложность, так как стадию «гимнодиниум» в жизненном цикле имеют, помимо атекальных, многие паразитические, а также свободноживущие панцирные виды. При идентификации гимнодиниоидов на уровне рода, помимо формы тела, решающее значение часто имело положение, направление и форма пояска и борозды, а также длина последних; степень расхождения концов пояска, соотношение верхней (эписомы) и нижней (гипосомы) частей тела. Однако этих признаков нередко было явно недостаточно. Особенно нелегкие таксономические проблемы возникали при идентификации видов двух крупнейших (явно сборных) родов Gymnodinium и Gyrodinium. Исследованиями последних лет ряда фототрофных динофлагеллят, с помощью методов фено – и геносистематики, было решено несколько актуальных из этих проблем (Daugbjerg et al., 2000; Hansen et al., 2000). Так, используя в качестве основного признака направление и форму так называемой апикальной бороздки (акробазы), на которую было обращено внимание еще в 1934 году (см. Biecheler, 1952, ей же принадлежит термин “acrobase”), род Gymnodinium был разделен на 4 рода: собственно Gymnodinium, Karenia G. Hansen et Moestrup, Karlodinium J. Larsen и Akashiwo G.Hansen et Moestrup. Род Gyrodinium был описан заново. Проблематичным остается довольно крупный и в большой степени искусственный род Amphidinium, однако для переописания его необходимо иметь гораздо большее количество данных, чем удалось получить до сих пор. К сожалению, подобные исследования возможны пока, главным образом, только с фототрофными (вероятно, не самыми многочисленными в вышеперечисленных крупных родах видами), относительно легко поддающимися культивированию, с помощью которого можно получить нужный для изучения объем материала.
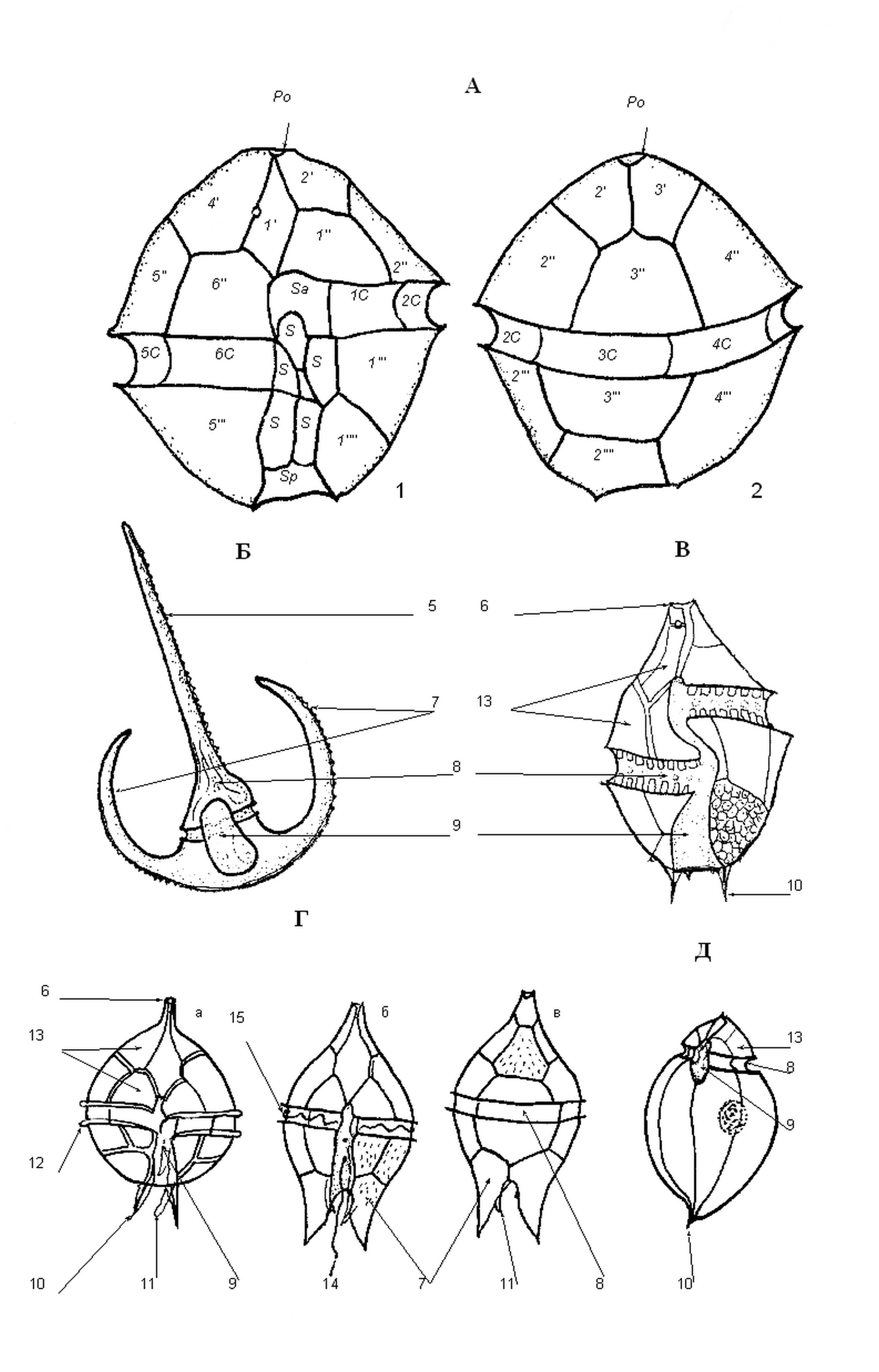 Рисунок 3. Общий вид и схема строения панцирных перидиней – гониаулакоидов и перидиниоидов: А – строение теки на примере рода Alexandrium с вентральной (1) и дорсальной (2) сторон клетки: Po – апикальная пора, или апикальный поровый комплекс; 3, 4 – текальные пластинки эпитеки и гипотеки; пластинки пояска (C) и борозды (S). Общий вид и строение панциря родов Ceratium (Б – вид спереди),Gonyaulax (В – вид спереди),Protoperidinium (Г: а, б – вид спереди, в – вид сзади), Oxytoxum (Д – вид спереди): 5 – апикальный рог, 6 – апикальный рог или вырост, 7 – антапикальные рога, 8 – поясок (поперечная борозда), 9 – борозда (собственно борозда, или продольная борозда), 10 – антапикальные шипы, 11 – бороздковая перепонка (краевой вырост, лист), 12 – поясковая мембрана, 13 – текальные пластинки, 14 – бороздковый жгутик, 15 – поясковый жгутик.
Рисунок 3. Общий вид и схема строения панцирных перидиней – гониаулакоидов и перидиниоидов: А – строение теки на примере рода Alexandrium с вентральной (1) и дорсальной (2) сторон клетки: Po – апикальная пора, или апикальный поровый комплекс; 3, 4 – текальные пластинки эпитеки и гипотеки; пластинки пояска (C) и борозды (S). Общий вид и строение панциря родов Ceratium (Б – вид спереди),Gonyaulax (В – вид спереди),Protoperidinium (Г: а, б – вид спереди, в – вид сзади), Oxytoxum (Д – вид спереди): 5 – апикальный рог, 6 – апикальный рог или вырост, 7 – антапикальные рога, 8 – поясок (поперечная борозда), 9 – борозда (собственно борозда, или продольная борозда), 10 – антапикальные шипы, 11 – бороздковая перепонка (краевой вырост, лист), 12 – поясковая мембрана, 13 – текальные пластинки, 14 – бороздковый жгутик, 15 – поясковый жгутик.
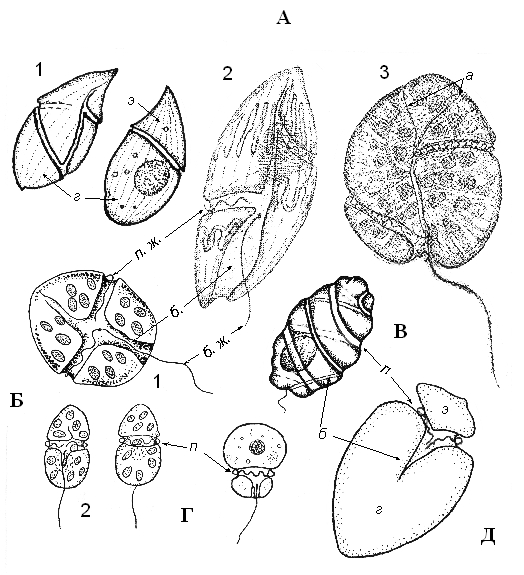
Рисунок 4. Основные типы строения беспанцирных перидиней – гимнодиниоидов. А (1-3) –Gyrodinium; Б (1,2) – Gymnodinium; В – Cochlodinium; Г – Katodinium; Д – Amphidinium: а – акробаза, б – борозда, п – поясок, э – эписома, г – гипосома, п.ж. – поясковый и бороздковый (б.ж.) жгутики (А (2, 3) – по: Biecheler, 1952).